 | Background - Place Cells Model - Boundary Vector Cells Navigation Spatial Memory Back to my Homepage For Potential Graduate Students |
| Background - Place Cells Model - Boundary Vector Cells Navigation Spatial Memory Back to my Homepage For Potential Graduate Students |
Cells in the rat's hippocampus fire when the animal is in a particular part of its environment (see Figure 1). For instance, if an animal is foraging in a rectangular box, a cell might fire whenever it is in the north-east corner. These neurons are called "place cells", and the region of the environment in which a given cell fires is called its place field. Place cells literally encode location in the sense that by looking at the firing of a population of place cells, one can tell to within a few centimeters where the animal is.
A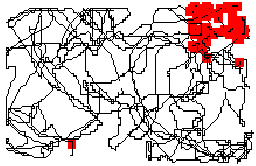
| B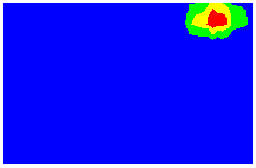
|
The place cell representation of space is different from other spatial representations in the brain, many of which can be thought of as capturing the relationship between individual stimuli and the body or some part of it (i.e., egocentric representations). When multiple cues are present, place cell firing does not depend on any single cue (subsets of cues can be removed, and the place fields are unaffected) but rather on the configuration of spatial cues. Place cell firing is also independent of the animal's current heading. O'Keefe and Nadel proposed that this special type of "allocentric" representation could provide the basis for a cognitive map, a representation that provides the information needed to locate oneself with respect to the world and to find one's way about.
Evidence from experiments which look at animal behaviour is consistent with the idea that the hippocampus is critical for spatial memory and navigation. For instance, in the "Morris Water Maze" paradigm, rats with damage to the hippocampus take longer to find a platform hidden beneath the surface of a pool of opaque liquid, and when the platform is removed, they fail to search for it in the right place. This kind of task, which involves remembering a hidden location with reference to several spatial cues is particularly sensitive to hippocampal damage. Other tasks which can be solved with reference to a single cue are unaffected. It seems reasonable to imagine that representation of location provided by place cell plays a part in this kind of task.
But how is the place cell representation formed? An important clue comes from experiments showing that a given place field will tend to maintain a fixed-distance to part of the boundary when the size or shape of the environment changes: place fields typically "stick" to part of the boundary (see Figure 2).
A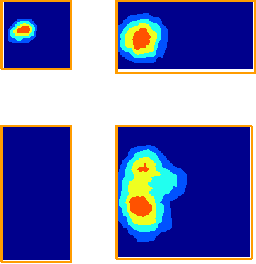 | B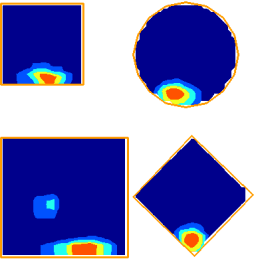 |
With Neil Burgess and colleagues, I developed a model (see Figure 3) of place cell firing based on the idea that the inputs to place cells respond to boundaries at particular distances and compass directions. Patterns of place cell firing across different environments can be explained assuming inputs from a small number of these hypothetical "boundary vector cells". This very simple model can also predict the firing field of a given cell in a new environment.
A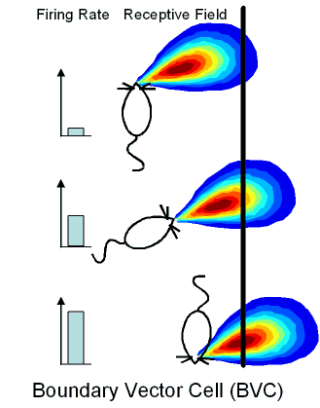 |
B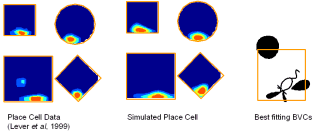 C 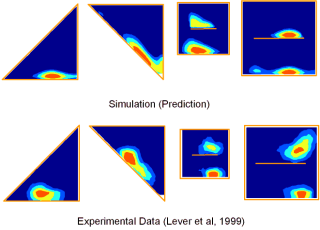
|
Cells with properties similar to place cells have recently been discovered in the human hippocampus, so one might be expect it to play a similar role in spatial memory and navigation. However, the idea that the hippocampus acts as a cognitive map in humans is controversial, since it is clear that in our species it is also involved in non-spatial aspects of memory. In particular, damage to the hippocampus results in a loss of the ability to form new episodic memories (memories for personally experienced events and their context). In this form of amnesia some other forms of memory (for instance memory for skills such as the ability to ride a bike or play the piano) are generally unaffected. It has been hypothesized that the role of the hippocampus in episodic memory is an extension of the spatial function seen in animals, or conversely that the spatial functions seen in animals are just one manifestation of a more general mnemonic function. The relationship between the spatial and non-spatial functions of the human hippocampus is intriguing and the subject of ongoing research. For instance we have found that, for a patient with damage to the hippocampus, difficulty in remembering locations viewed from a novel perspective (allocentric spatial memory) is associated with problems in recalling the context in which an event occurred (episodic memory), but not the content of the event itself.
A
|
B i) ii)
ii)
|
However this episodic memory function is related to its spatial function, it is clear that the hippocampus remains critical to some forms of spatial behaviour in humans, especially navigation in large scale environments. For instance, one of our own studies showed that activity in the hippocampus is correlated with performance on a virtual navigation task. Subjects in an fMRI scanner found their way between locations in one of two virtual towns. In one town subjects had to choose new direct paths that they had not previously followed. In the other they had to follow a fixed route that they had previously practiced repeatedly.
A
|
B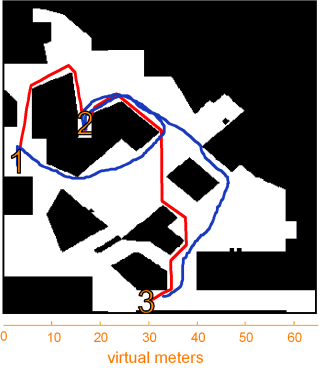
|
When subjects were finding new direct paths we found a correlation between individual performance and activity in the hippocampus: the more direct the path a subject took, the more activity was seen in the hippocampus. In another part of the brain, the caudate nucleus, better navigators showed more activation when following a fixed route. In animals the caudate is associated with the learning fixed patterns of behaviour (for example turning left, then turning right). This type of learning, which does not depend on the hippocampus, would be useful when following the same route repeatedly - it could act a little like an autopilot, taking control of behaviour and freeing up other parts of the brain to be used for more complex tasks. However an autopilot would not be useful for planning new direct routes, where a cognitive map would be more useful.
A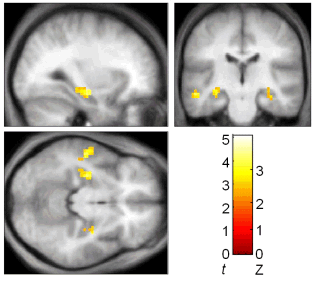
|
B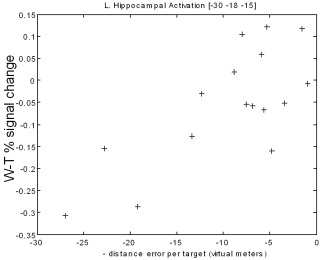
|
C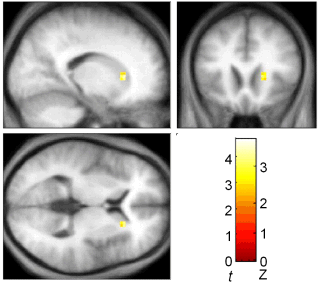
|
D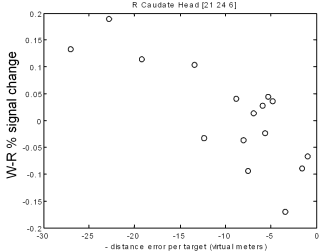
|
One interpretation of our findings is that the two brain regions compete to control navigation behaviour. Good navigators characteristically choose the "right" neural representation for the task at hand (i.e., the hippocampal "cognitive map" when finding their way, the caudate "autopilot" when following a familiar route). Poor navigators by contrast often activate the wrong system.
If the hippocampus acts as a cognitive map, then our model place cell model suggests that the map will be distorted in a predictable way if the geometry of the environment is manipulated. We tested this by asking subjects to remember the location of a golf flag in a virtual reality arena. The subjects were temporarily removed from the arena, and then put back at a random location. In the meantime we changed the shape and size of the arena. We found that when subjects were asked to indicate where the golf flag had been (by marking the spot with a virtual traffic cone), their responses followed a predictable pattern which could be explained by our place cell model.
A
|
B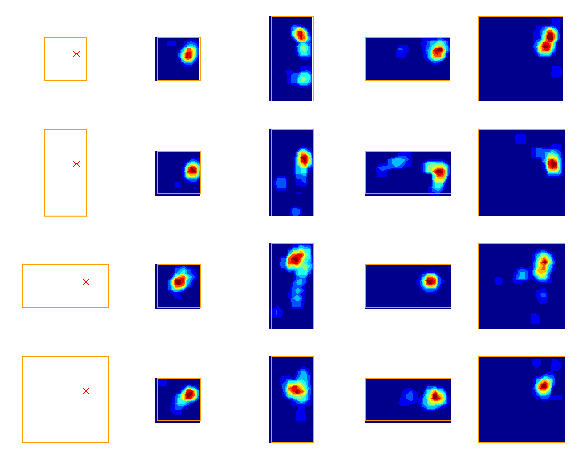
|
C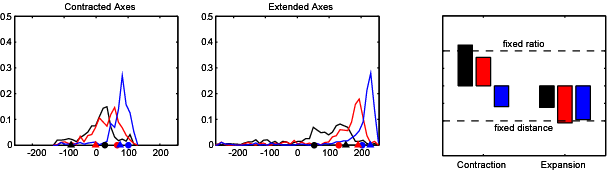
|
D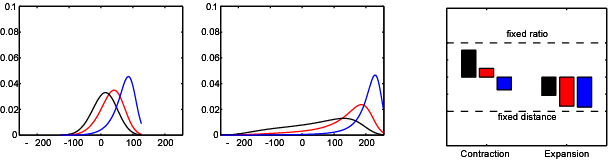
|